CORPO
Dalle reti fluviali al metabolismo degli organismi viventi
di Amos Maritan
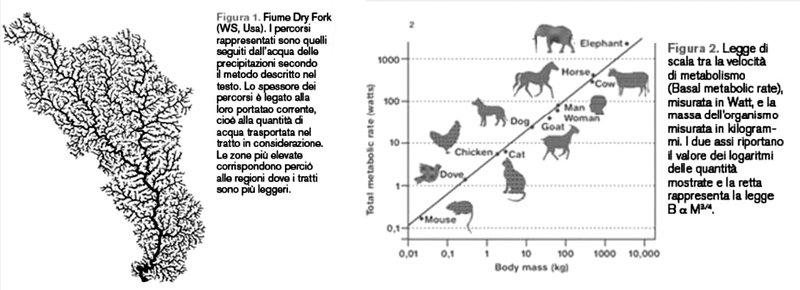
Recentemente si è scoperto che le proprietà della rete di trasporto dei nutrimenti – il sistema venoso – negli organismi viventi (J. Banavar, A. Maritan, A. Rinaldo, Size and form in efficient transportation networks, in «Nature», 1999), mammiferi compresi, è responsabile del legame che c’è tra la massa corporea, M, e il metabolismo basale, B. Il nostro metabolismo basale misura, per esempio, quanto calore dissipiamo ogni ora, oppure quanto ossigeno consumiamo, sempre ogni ora, in condizioni di riposo. Lo stesso approccio usato per comprendere l’ottimalità intrinseca delle reti fluviali e le loro proprietà (Banavar et al., cit.) ha permesso di fornire una spiegazione di uno dei più grandi puzzle della macro-ecologia moderna, formulato da Kleiber nel 1932. La legge di Kleiber (M. Kleiber, The Fire of Life, 1975) afferma che gli organismi viventi, da quelli formati solamente da alcune cellule a quelli più grandi come la balena blu (Banavar, Maritan, Rinaldo, cit.), hanno un metabolismo basale che obbedisce alla legge di potenza B = C M3/4 (figura 2). Il puzzle nasceva dal fatto che un semplice ragionamento geometrico portava a un esponente 2/3 invece che 3/4 (B è proporzionale all’area della superficie del corpo attraverso cui avviene la dissipazione di calore e questa, a sua volta, si comporta come M2/3). Questa legge ha talmente tante conseguenze che è impossibile elencarle in questa breve nota. Ne menzioniamo solo alcune. La durata della vita di un organismo di massa M è dell’ordine di M1/4 e, come conseguenza, un elefante di 6 tonnellate vive circa 5 volte più di un cane di 10 kg. Il battito cardiaco dell’elefante è 5 volte più basso di quello del cane, e il cane dorme molto più dell’elefante, ecc. La biodiversità in una foresta incontaminata si deduce dalla legge di Kleiber. Per esempio, il numero di piante con un certo diametro, D, presenti nella foresta è proporzionale a 1/ D2, un’altra legge di potenza. È sorprendente che una legge così semplice e con così tante conseguenze abbia trovato una spiegazione solo di recente (Banavar, Maritan, Rinaldo, cit.) e tramite l’analogia con le reti fluviali.
I bacini fluviali sono un bellissimo esempio di strutture frattali che hanno subito un’evoluzione auto-organizzandosi in geometrie tanto più stabili quanto più variabili sono state le condizioni geologiche e climatiche in cui essi si sono sviluppati (I. Rodriguez Iturbe, A. Rinaldo, Fractal River Basins: Chance and Self-Organization, 1996). Lo scopo di questa nota è quello di spiegare l’origine di queste forme frattali e la loro relazione con le leggi che regolano la variazione del metabolismo degli organismi viventi in funzione della loro massa corporea.
Le forme dei bacini fluviali possono essere visualizzate con le mappe di elevazione digitale. Una mappa di elevazione digitale, ottenuta con tecniche di altimetria laser (remote sensing), non è altro che il campo delle altezze del terreno definite su una ‘griglia’ quadrata, in ogni nodo della quale è definita l’altezza media del terreno. Il percorso seguito dall’acqua è quello della ‘discesa più rapida’: l’acqua dovuta alle precipitazioni in una certa area della griglia fluisce successivamente nell’area limitrofa con l’altezza più bassa tra tutte le aree vicine. Il processo continua finché l’acqua raggiunge la pianura, l’uscita del bacino.
La figura 1 mostra la forma della rete fluviale corrispondente a un fiume del Nordamerica, il Dry Fork, ottenuta con il metodo appena descritto. Si noti che la rete dei percorsi seguiti dall’acqua non ha circuiti chiusi: questo fatto viene ricordato dicendo che la rete è un ‘albero’. È evidente dalla figura il carattere ‘frattale’ o auto-simile del bacino: ogni sotto-bacino assomiglia a una copia rimpicciolita dell’intero bacino, ovvero ogni sua parte è ‘simile’ al tutto. Nella sua discesa l’acqua perde energia potenziale gravitazionale. Una parte di questa energia va in energia cinetica necessaria per il deflusso dell’acqua stessa dal bacino, mentre la rimanente parte viene dissipata.
I dati delle osservazioni su reti fluviali in vari punti della Terra e gli studi di modelli matematici per le reti di trasporto hanno portato alla conclusione che le forme dei bacini osservati corrispondono a una evoluzione che ‘tenta’ di minimizzare l’energia dissipata (Rodriguez Iturbe, Rinaldo, cit.; J. Banavar et al., Scaling, Optimality and Landscape Evolution, in «Journal of Statistical Physics», 2001). In altre parole, tra tutte le reti di trasporto possibili quella, per esempio, mostrata in figura 1 dissipa meno energia di un’altra rete compatibile con i vari vincoli geologici presenti nel bacino. Inondazioni, frane ecc. hanno contribuito all’evoluzione della rete, che è tanto più stabile, od ottima, quanto meno dissipa energia.
Esperimenti numerici e argomenti teorici mostrano che le reti fluviali dei bacini di tutta la Terra sono caratterizzate dalle cosiddette leggi di potenza. Descriviamo la più importante per dare un’idea di come si possa quantificare l’osservazione fatta sopra a proposito dell’auto-similarità della rete fluviale. Ogni tratto del fiume di figura 1 è caratterizzabile dalla quantità di acqua, J, che può trasportare, diciamo in un giorno. La percentuale di tratti di fiume con una corrente più grande di una certa J, presenti nel bacino fluviale, ha una legge estremamente semplice e valida per tutti i bacini, cioè P(J) = C J-a dove ‘a’ è un esponente che varia tra 0,42 e 0,46 per i vari bacini fluviali, mentre C è una semplice costante. Questo tipo di legge si chiama legge di potenza. Le strutture frattali possono essere caratterizzate da vari tipi di leggi di potenza. L’importanza di tali leggi sta nel fatto che l’esponente della legge è una quantità ‘emergente’, cioè non dipende dai dettagli del fenomeno in questione. Infatti, come appena osservato, indipendentemente dal fatto che si tratti di fiumi in Cina o America, l’esponente ‘a’ è lo stesso (a parte le incertezze legate alle misure).